Results
A multivariate regression tree (MRT) was used to examine the relationship between the dominant vegetation type and sex of bear associated with each site (predictor variables), and the most frequently observed species (10 or more observations) (Figure 10). This tree shows no splits using sex of bear and although it does show divisions based on dominant vegetation type, the error value of 0.941 is far too high to draw any assumptions from this tree. One aspect of this MRT to note is the relationship between the CHSP (chipping sparrow) and the SHRUB_MIXED (shrub and mixed forest) branch of the tree. Again, given the error value, I wouldn't draw any strong conclusions from this MRT, but this may perhaps indicate a potential relationship between this vegetation type and the chipping sparrow.
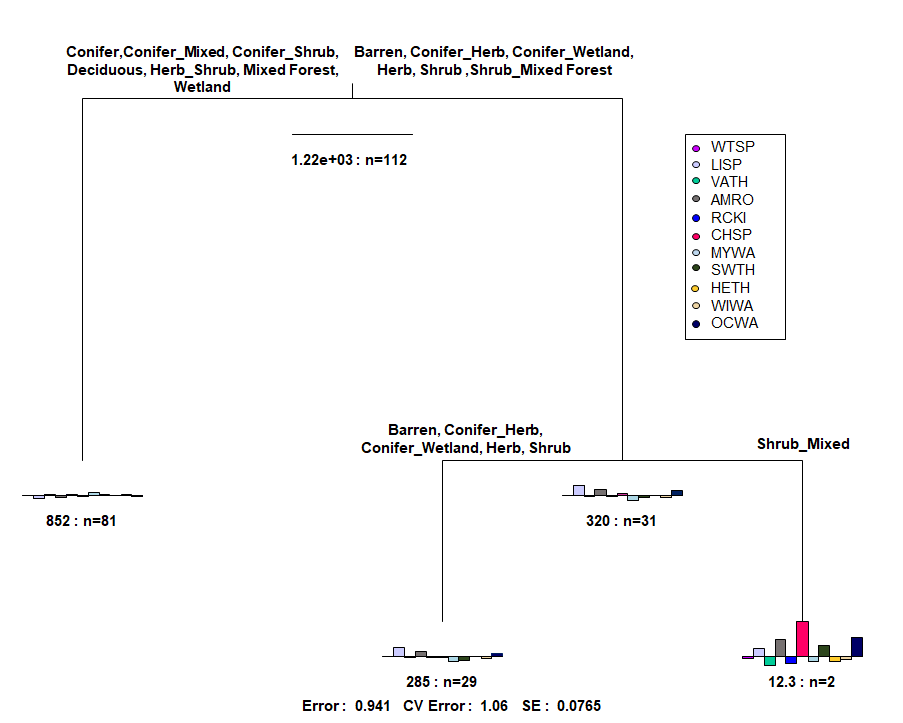
Figure 10. Multivariate Regression Tree of songbird species with 10 or more observations, with dominant vegetation type and sex of bear (male, female, or no bear) as predictor variables.
We then used a Permutational Multivariate Analysis of Variance (PERMANOVA) to look for any relationships between our two predictor variables and the songbird species observed. For both PERMANOVAs, Bray-Curtis was used as the distance measure.
Firstly, no significant difference was present in the songbird species observed between the three different sex of bear (male, female, or no bear) site types (Table 1).
Firstly, no significant difference was present in the songbird species observed between the three different sex of bear (male, female, or no bear) site types (Table 1).
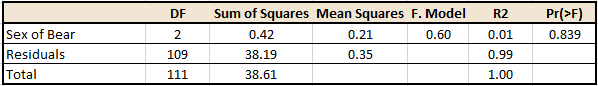
Table 1. Output from the PERMANOVA examining the relationship between the sex of the bear and the songbird species observed at each site.
Secondly, a significant relationship was found (p=0.007) between the dominant vegetation types, based on the songbird species observed (Table 2).
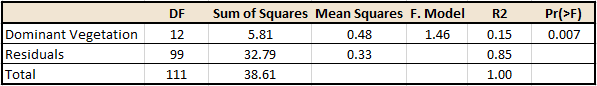
Table 2. Output from the PERMANOVA examining the relationship between the dominant vegetation type and the bird species observed at each site.
Following the significant result found in Table 2, pairwise PERMANOVAs (also using Bray-Curtis as distance) were done for each possible 77 pairings of vegetation types. The p-values from these tests were then adjusted using the Holm method. The output from these tests (Table 3) do not indicate any significant differences between the dominant vegetation types. This does contradict the PERMANOVA results found in Table 2. One possible explanation for this discrepancy could be a type 1 error in the original PERMANOVA.
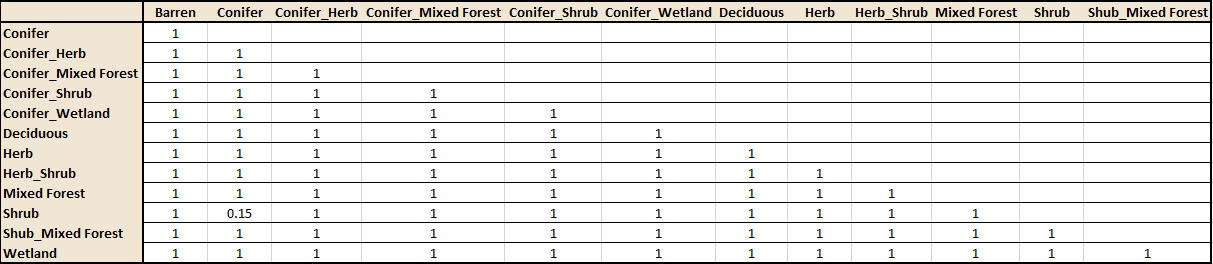
Table 3. Output from the PERMANOVA pairwise comparisons of songbird species based on dominant vegetation type, with p-values adjusted using the Holm method.
Discussion
 Figure 11. Illustration of radius of micro-site and therefore vegetation surveys with radius of ARU recording distance potential overlain.
Figure 11. Illustration of radius of micro-site and therefore vegetation surveys with radius of ARU recording distance potential overlain.
The results from this study generally conclude that 1) bird species values are not impacted by the vegetation at our sites and 2) bird species presence (and number of observations) is not significantly impacted by bear presence (based on our analyses of sex of the bear associated with each site).
Firstly, the result of no relationship between the vegetation type and the bird species is unusual given the known relationship of bird species to different vegetation types, such as those observed by Hobson and Bayne (2000). One potential explanation for this result could be the scale at which we measured the vegetation. As we were wanting to look at bear micro-sites, we chose a 15 m radius to represent the small scale at which the bear would have selected the site and measured the vegetation within this radius. However, our means of detecting birds (autonomous recording units) are capable of recording audio far beyond this radius (Yip et al 2017); therefore, our vegetation measurements reflect only a portion of the habitat that the birds we hear on the recordings are truly occupying (as illustrated in Figure 11).
Secondly, to address our conclusion of a non-relationship between bear use of sites and the songbird species observed, as is discussed in more detail above, the autonomous recording units are not truly providing a recording representative of the species present within the boundaries of our small micro-site - not allowing using to differentiate between what is within the radius and what is without.
From this study, I can't conclude that grizzly bears are an appropriate surrogate for songbird diversity; however, I believe that the issue of the scale of measurement for the bear and vegetation variables versus the scale of measurement of songbirds doesn't allow this data to answer the question of whether grizzly bears are capable of acting as a surrogate species.
To further examine this surrogacy question, I would recommend looking at a scale where the bird data and the relevant predictor variables are more aligned in their area of measurement. Regardless of the results of this study, I would recommend examining different sizes of areas occupied by bears (and the bird species within them), such as home ranges, as opposed to only micro-sites, as the size of the area managed can impact how well species are conserved (Higa et al 2016).
Firstly, the result of no relationship between the vegetation type and the bird species is unusual given the known relationship of bird species to different vegetation types, such as those observed by Hobson and Bayne (2000). One potential explanation for this result could be the scale at which we measured the vegetation. As we were wanting to look at bear micro-sites, we chose a 15 m radius to represent the small scale at which the bear would have selected the site and measured the vegetation within this radius. However, our means of detecting birds (autonomous recording units) are capable of recording audio far beyond this radius (Yip et al 2017); therefore, our vegetation measurements reflect only a portion of the habitat that the birds we hear on the recordings are truly occupying (as illustrated in Figure 11).
Secondly, to address our conclusion of a non-relationship between bear use of sites and the songbird species observed, as is discussed in more detail above, the autonomous recording units are not truly providing a recording representative of the species present within the boundaries of our small micro-site - not allowing using to differentiate between what is within the radius and what is without.
From this study, I can't conclude that grizzly bears are an appropriate surrogate for songbird diversity; however, I believe that the issue of the scale of measurement for the bear and vegetation variables versus the scale of measurement of songbirds doesn't allow this data to answer the question of whether grizzly bears are capable of acting as a surrogate species.
To further examine this surrogacy question, I would recommend looking at a scale where the bird data and the relevant predictor variables are more aligned in their area of measurement. Regardless of the results of this study, I would recommend examining different sizes of areas occupied by bears (and the bird species within them), such as home ranges, as opposed to only micro-sites, as the size of the area managed can impact how well species are conserved (Higa et al 2016).
References
Higa, M., Yamaura, Y., Senzaki, M., Koizumi, I., Takenaka, T., Masatomi, Y., Momose, K. (2016). Scale Dependency of two endangered charismatic species as biodiversity surrogates. Biodiversity and Conservation, 25, 1829-1841.
Hobson, K.A. and Bayne, E. (2000). Breeding Bird Communities in a Boreal Forest of Western Canada: Consequence of "Unmixing" the Mixedwoods. The Condor, 102, 759-769.
Yip, D., Bayne, E.M., Sólymos, P., Campbell, J., Proppe, D. (2017). Sound attenuation in forest and roadside environments: Implications for avian point count surveys. The Condor: Ecological Applications, 119, 73-84.
Higa, M., Yamaura, Y., Senzaki, M., Koizumi, I., Takenaka, T., Masatomi, Y., Momose, K. (2016). Scale Dependency of two endangered charismatic species as biodiversity surrogates. Biodiversity and Conservation, 25, 1829-1841.
Hobson, K.A. and Bayne, E. (2000). Breeding Bird Communities in a Boreal Forest of Western Canada: Consequence of "Unmixing" the Mixedwoods. The Condor, 102, 759-769.
Yip, D., Bayne, E.M., Sólymos, P., Campbell, J., Proppe, D. (2017). Sound attenuation in forest and roadside environments: Implications for avian point count surveys. The Condor: Ecological Applications, 119, 73-84.
